
Тельце барра и синдром дауна

Решение задачи 1
Кариотип 1
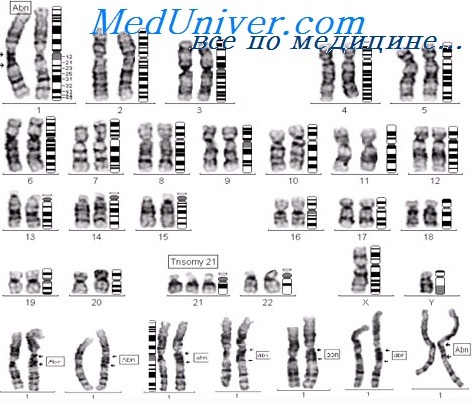
47, ху +21 патология, синдром Дауна, трисомия по 21 паре хромосом
Кариотип 2
47, ху +13 патология, синдром Патау, трисомия по 13 паре хромосом
Кариотип 3
47, Х0 патология, синдром Шерешевского –Тернера, моносомия трисомия по 23 паре хромосом
Кариотип 4
47, ХХУ патология, синдром Клайнфельтера, трисомия по 23 паре хромосом
Кариотип 5
47, ху +18 патология, синдром Дауна, трисомия по 21 паре хромосом
Задача №2 . Решение задач на дисбаланс половых хромосом и определение
количества телец Барра
Определите количество полового хроматина у людей с дисбалансом половых хромосом, укажите число хромосом, синдром:
а)у мужчины ХХУ, ХХХУ, ХХХХУ ; б) у женщин ХО, ХХХ.
Решение задачи 2
А) Количество полового хроматина у мужчины с дисбалансом половых хромосом ХХУ-1 тельце Барра, синдром Клайнфельтера
ХХХУ – 2 тельца Барра, синдром Клайнфельтера
ХХХХУ – 3 тельца Бара, синдром Клайнфельтера
Б) Количество полового хроматина у женщины с дисбалансом половых хромосом составляет: при
ХО – ни одного, синдром Шерешевского –Тернера
ХХХ-2 тельца Баррра, синдром трипло-Х
Решение задач на нахождение гена на картах дифференциально окрашенных хромосом
Задача №3 Расшифруйте условные обозначения и отметьте стрелками соответствующие сегменты хромосом на рисунках:
а)3р13; б)3g29; в)3р26; г) 4р15; д)4g26; е) 11 р 14; ж)11g 26
Решение задачи
а) третий сегмент первого района короткого плеча третьей хромосомы;
б) девятый сегмент второго района длинного плеча третьей хромосомы;
в) шестой сегмент второго района длинного плеча третьей хромосомы;
г) пятый сегмент первого района короткого плеча четвертой хромосомы;
д) шестой сегмент второго района длинного плеча четвертой хромосомы;
е) четвертый сегмент первого района короткого плеча одиннадцатой хромосомы;
ж) шестой сегмент второго района длинного плеча одиннадцатой хромосомы.
Задача №4
С помощью современных молекулярно-генетических методов в ДНК обнаружены:
а) локус гена WHCR – ген синдрома Вольфа (низкий рост, микроцефалия, судорожные припадки, множественные аномалии развития), имеющего координаты 4 р16;
б) локус гена NF – ген нейрофиброматоза 2 типа, имеющего координаты 22 g12
в) локус гена CLS – ген синдрома Коффина Лоури (умственная отсталость, «куриная грудь», «нос боксера», гипертелоризм), Хр22;
г) локус гена GEY – зеленый/синий цвет глаз, имеющего координаты 19 р13
д) локус гена NPC – болезнь Нимана-Пика, имеющего координаты 18 g 11- g 12;
е)локус гена онкогена ras, имеющего координаты 3р25;
ж)локус гена ОFS1, CL – расщепление губы с расщеплением или без расщепления неба, 6р24
з) локус гена NF – ген нейрофиброматоза 2 типа, имеющего координаты 22 g12
Решение задачи 4
.
Этап: Подведение итогов, домашнее задание
1. Повторить к следующему занятию учебный материал по генеалогическому методу;
2. Дать характеристику кариотиограмме.
 А) Б)
А) Б)
 В)
В)
Г)

Д)
Методические указания для студентов к практической работе №1.
Цель: Научиться отличать нормативный кариотип женщины и мужчины от патологического
по кариограммам и количеству телец Барра
Оснащение занятия: рисунки
1. “Видимое строение хромосом”.
2. “Хромосомный набор мужчины и женщины”.
3. “Схематическое изображение дифференциальной окраски хромосом человека по G-методу”.
4. Рисунок «Ядра клеток буккального эпителия».
Ход работы:
1. Изучите по схемам видимое строение хромосом (рисунок 1, 2) и зарисуйте особенности их морфологии.

2. Изучите классификацию и номенклатуру равномерно окрашенных хромосом (рисунок 3,4). Оответьте на вопросы:
– Какая классификация хромосом принята в качестве основной?
– В чем заключаются возможности Денверской классификации хромосом?
– По какому принципу разделены хромосомы в этой классификации?
Классификация и номенклатура равномерно окрашенных хромосом человека впервые были приняты в 1960 году в г. Денвере. Согласно классификации все хромосомы человека разделены на 7 групп, расположенных в порядке уменьшения их длины, и обозначаются буквами латинского алфавита от A до G. Все пары хромосом стали нумеровать арабскими цифрами.
(1- 3) – самые большие хромосомы; 1 и 3-я – метацентрические, 2-я – субметацентрические.
4 и 5-я)- крупные субметацентрические хомосомы.
Группа С (6-12-я и Х-хромосома)- субметацентрические хромосомы среднего размера.
Группа D(13-15-я)- акроцентрические хромосомы средних размеров.
Группа Е (16-18-я)- маленькие субметацентрические хромосмы.
Группа F(19-20-я)- самые маленькие метацентрические хромосомы.
Группа G((21,22-я и Y) –самые маленькие акроцентрические хромосомы.
Предложенная классификация позволяла четко различать хромосомы, принадлежащие к разным группам.
3. Выполните задание
Задача 1.
Среди предложенных кариотипов (рисунок 6 , кариотипы 1-5), укажи аномальные, запиши их кариотип и определи вид хромосомной анеуплоидии: нулесомия, моносомия, трисомия, если возможно – назови синдром.

4. Изучите принципы определения определение полового хроматина
Хроматин клеточного ядра подразделяется на два основных типа на эу – и гетеро хроматин. Это наследственный материал различной степени спирализации и упаковки белками различной степени конденсации.
Эухроматин (от греч. еu — полностью и сhгоmа – цвет) в метафазных хромосомах виден в виде светлых полос.
В эухроматине находятся структурные активные уникальные гены, которые контролируют развитие признаков организма. Эухроматин менее плотно упакован и доступен для ферментов РНК-полимераз, обеспечивающих синтез и-РНК, а затем синтез белков.
Гетерохроматин выявляется в метафазных хромосомах при дифференциальном окрашивании в виде темных полос различных размеров, состоящих из конденсированной (спирализованной) плотно упакованной молекулы ДНК. Даже в интерфазном ядре гетерохроматин в виде глыбок хорошо виден в световой микроскоп Чаще всего он расположен вокруг ядрышка и около ядерной оболочки. Переписывания информации и-РНК с данных участков не происходит. Эти гены неактивны.
Структурный и факультативный гетерохроматин
Структурный гетерохроматин в интерфазном ядре спирализирован, плотно упакован в метафазных хромосомах постоянно обнаруживается вокруг центромеры во всех 46 хромосомах (составляет около 13 % от генома). Расположение темных полос для каждой пары хромосом строго индивидуально. Функция структурного гетерохроматина в целом пока неясна.
Факультативный гетерохроматин появляется в интерфазном ядре не всегда. Это спирализованный эухроматин. В метафазных хромосомах факультативный гетерохроматин не обнаруживают. Например, в ядрах клеток женщин в диплоидном наборе имеется две Х-хромосомы, одна из которых полностью инактивирована (спирализована, плотно упакована) уже на ранних этапах эмбрионального развития и видна в виде глыбки гетерохроматина, прикрепленного к оболочке ядра. Благодаря этому женские и мужские организмы уравновешиваются по количеству функционирующих генов, сцепленных с полом, так как у мужчин одна-X-хромосома и одна доза генов Х-хромосомы. Инактивированная Х-хромосома называется половым хроматином или тельцем Бара (рисунок 5).

Половой хроматин обычно определяют путем анализа эпителиальных клеток в соскобе слизистой оболочки щеки. Отсутствие тельца Барра у женщин свидетельствует о хромосомном заболевании – синдроме Шерешевского-Тернера, присутствие у мужчин тельца Барра свидетельствует о наследственном заболевании – синдроме Клайнфельтера (кариотип 47, XXV).
5. Выполните задание
Задача 2
Определите количество полового хроматина у людей с дисбалансом половых хромосом, укажите число хромосом, синдром:
а)у мужчины ХХУ, ХХХУ, ХХХХУ ; б) у женщин ХО, ХХХ.
6. Изучите методы цитологического окрашивания хромосом
Наиболее подходящей фазой для исследования хромосом является метафаза митоза. Для изучения хромосом чаще используют препараты кратковременной культуры крови, полученные через 48 -72 ч после взятия крови. При приготовлении препаратов хромосом к культуре клеток добавляют колхицин, который разрушаем веретено деления и останавливает деление клетки в метафазе. Затем клетки обрабатывают гипотоническим раствором, после чего их фиксируют и окрашивают.
А) Полное окрашивание хромосом. Для окраски хромосом чаще используют краситель Романовского—Гимзы, 2% ацеткармин или 2 % ацетарсеин. Они окрашивают хромосомы целиком, равномерно (ругинный метод) и могут быть использованы для выявления численных аномалий хромосом человека (45, 47 и т. д.).
Б) Дифференцированное окрашивание хромосом.
Для получения детальной картины структуры хромосом используют различные способы дифференциального окрашивания. Один из них – G-метод: по длине хромосомы выявляется ряд окрашенных и неокрашенных полос. Чередование этих полос и их размеры строго индивидуальны и постоянны для каждой пары гомологичных хромосом. Например, хромосомы 13, 14, 15-й пар трудно отличить при равномерной окраске, а при дифференциальной – рисунок исчерченности (чередование и размер темных и светлых полос) неодинаков ( рисунок 6).
6. Выполните задания
Задача 3
Расшифруйте условные обозначения и отметьте стрелками соответствующие сегменты хромосом на рисунках:
а)3р13; б)3g29; в)3р26; г) 4р15; д)4g26; е) 11 р 14; ж)11g 26
Задача 4
С помощью современных молекулярно-генетических методов в ДНК обнаружены:
а) локус гена WHCR – ген синдрома Вольфа (низкий рост, микроцефалия, судорожные припадки, множественные аномалии развития), имеющего координаты 4 р16;
б) локус гена NF – ген нейрофиброматоза 2 типа, имеющего координаты 22 g12
в) локус гена CLS – ген синдрома Коффина Лоури (умственная отсталость, «куриная грудь», «нос боксера», гипертелоризм), Хр22;
г) локус гена GEY – зеленый/синий цвет глаз, имеющего координаты 19 р13
д) локус гена NPC – болезнь Нимана-Пика, имеющего координаты 18 g 11- g 12;
е)локус гена онкогена ras, имеющего координаты 3р25;
ж)локус гена ОFS1, CL – расщепление губы с расщеплением или без расщепления неба, 6р24
з) локус гена NF – ген нейрофиброматоза 2 типа, имеющего координаты 22 g12
Из за большого объема этот материал размещен на нескольких страницах: 1 2 3 4 5 |
Источник
Половой хроматин. Тельца Барра
Х-хроматин (тельце Барра) представляет собой хромоцентр величиной около 1 мкм, красящийся всеми основными ядерными красителями более интенсивно, чем остальные хроматиновые структуры ядра. Фельген-положительная реакция свидетельствует о большой концентрации в нем ДНК.
Локализация Х-хроматина в ядре различна. В большинстве тканей он находится на внутренней поверхности ядерной оболочки и может иметь треугольную, плоско-выпуклую, трапециевидную, U-образную или гантелевидную форму. Иногда Х-хроматин имеет вид утолщения или зубца ядерной мембраны, соединенного с ядрышком тонкой хроматиновой нитью. В веретеновидных и палочковидных ядрах Х-хроматин располагается на одном из полюсов ядра.
Реже Х-хроматин располагается на ядрышке или в нуклеоплазме, при этой локализации он обладает сферической формой и трудно отличим от других хромоцентров, имеющих такой же размер, но неспецифических для пола. Поэтому в целях диагностики половой принадлежности клеток большинство исследователей учитывают хромоцентры, расположенные только у ядерной мембраны.
Положение Х-хроматина может меняться в одних и тех же клетках в зависимости от их функционального состояния, а также в процессе онтогенеза.
Х-хроматин обнаружен в клетках различных тканей у многих млекопитающих; у грызунов (хомяки, крысы, мыши, морские свинки) хроматиновые структуры ядер представлены большим количеством хромоцентров, затрудняющих выявление Х-хроматина. У человека половые различия в строении ядер установлены практически во всех тканях и органах.
Происхождение Х-хроматина. В процессе клеточного цикла хромосомы претерпевают закономерные преобразования, которые состоят в спирализации и деспирализации хромосом и их репродукции. В интерфазе максимально деспирализованные хромосомы образуют ядро с относительно гомогенным содержимым. Репродукция (синтез ДНК) хромосом происходит только в деспирализованном состоянии в период S-интерфазы.
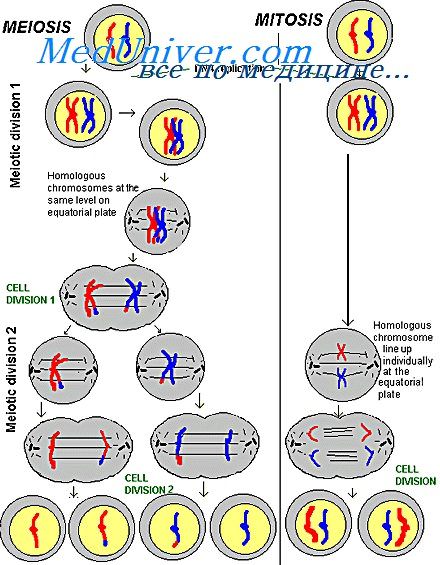
Спирализуясь, хромосомы вступают в профазу митоза и достигают наибольшей спирализации в метафазе митоза и мейоза. При этом они обладают минимальной специфической активностью. Вместе с тем установлено, что хромосомы всегда неравномерно спирализованы по длине и разделяются на гетерохроматические и эухроматические районы. Морфологически эти районы различаются по интенсивности окраски и структурной организации.
Эухроматические районы в интерфазном ядре деспирализуются, в то время как гетерохроматические имеют тенденцию оставаться в спирализованном компактном состоянии в виде хромоцентров с высоким содержанием ДНК. Спирализованность гетерохроматических районов сопровождается неактивным состоянием генов, содержащихся в них. Эта особенность свойственна также некоторым эухроматнческим районам с высоко функционально дифференцированными генами. Будучи спирализованными в стадии интерфазного ядра, эухроматические районы становятся также генетически неактивными.
Гетерохроматизация — универсальный механизм генетической инактивации хромосомных участков независимо от того, относятся ли они к гетерохроматическим или к эухроматнческим районам. Следовательно, хромоцентры, обнаруживаемые в интерфазном ядре, могут быть образованы как гетерохроматином, так и эухроматином. Одним из таких хромоцентров является Х-хроматин.
Еще Ваrr и Bertram высказали предположение о связи феномена Х-хроматина с Х-хромосомами. С тех пор Х-хромосомная природа Х-хроматина подтверждена и уточнена данными многочисленных исследователей.
Х-хроматин образован одной из Х-хромосом женской клетки, находящейся в гетерохроматизированном состоянии. Будучи спирализованной, эта хромосома генетически неактивна. В разных клетках сомы у женских особей, по принципу случайности, Х-хроматнн образует Х-хромосома, полученная либо от отца, либо от матери. Следовательно, клетки женского организма мозаичны по функции Х-хромосомы: в одних активна отцовская, в других материнская хромосома. Образование полового хроматина в женских клетках обусловлено генетически.
Это подтверждается тем, что в раннем периоде развития эмбриона человека, когда по виду гонад еще нельзя определить пол, яйцевые оболочки зародыша мужского пола не имеют Х-хроматина, несмотря на воздействие гормонов матери. У зародыша женского пола Х-хроматин появляется на 16-й день развития, когда в эмбрионе насчитывается 2500—5000 клеток.
– Также рекомендуем “Х-хроматин в клетках различных тканей. Наличие Х-хроматина в клетках”
Оглавление темы “Деление клеток. Половой хроматин”:
1. Обмен веществ в клетках. Раздражимость клеток
2. Телофаза и интерфаза митоза. Первое деление мейоза клеток
3. Второе деление мейоза. Амитоз и эндомитоз
4. Дифференциация клеток. Регенерация клеток
5. Способы регенерации клеток. Старение и гибель клетки
6. Удаление мертвых клеток из организма. Хромосомный пол
7. Половой хроматин. Тельца Барра
8. Х-хроматин в клетках различных тканей. Наличие Х-хроматина в клетках
9. Х-хроматин в лейкоцитах. Половые различия лейкоцитов
10. Половые виды лимфоцитов. Y-хроматин клеток организма
Источник
Генетика синдрома Дауна: кариотипКлинический диагноз синдрома Дауна обычно не представляет никаких трудностей. Тем не менее для подтверждения диагноза и предоставления базы для генетического консультирования необходимо кариотипирование. Хотя различия в конкретных вариантах кариотипа, ответственных за синдром Дауна, обычно имеют небольшое влияние на фенотип пациента, они существенны для определения риска повторения. Трисомия 21 при синдроме Дауна. Примерно у 95% всех пациентов с синдромом Дауна выявляют трисомию хромосомы 21, вызванную мейотическим нерасхождением 21 пары хромосом, как обсуждалось в предыдущей главе. Уже отмечено, что риск иметь ребенка с трисомией 21 увеличивается с возрастом матери, особенно после 30 лет. Мейотическая ошибка, ответственная за трисомию, обычно происходит в ходе материнского мейоза (около 90% случаев), преимущественно в первом делении, но около 10% случаев происходит в отцовском мейозе, обычно во втором делении. Робертсоновская транслокация при синдроме Дауна. Около 4% пациентов с синдромом Дауна имеют 46 хромосом, одна из которых — робертсоновская транслокация между хромосомой 21q и длинным плечом одной из других акроцентрических хромосом (обычно хромосомы 14 или 22). Транслоцированная хромосома заменяет одну из нормальных акроцентрических хромосом, и кариотип пациента с робертсоновской транслокацией между хромосомами 14 и 21 – 46,XX/XY,rob(14;21)(ql0;ql0),+21. Такая хромосома может также быть определена как der(14;21), на практике используют обе номенклатуры. В действительности пациенты с робертсоновской транслокацией, включающей хромосому 21, трисомны по генам, расположенным в длинном плече 21q. В отличие от стандартной трисомии 21, транслокационный синдром Дауна не показывает никакой связи с возрастом матери, но имеет сравнительно высокий риск повторения в семьях, если один из родителей, особенно мать, — носитель транслокации. По этой причине для точного генетического консультирования важно кариотипирование родителей и, возможно, других родственников.
Носители робертсоновской транслокации, включающей хромосомы 14 и 21, имеют только 45 хромосом; одна 14 и одна 21 отсутствуют и заменены транслоцированной хромосомой. Теоретически возможны шесть типов гамет, но три из них не могут привести к жизнеспособному потомству. Три типа гамет жизнеспособные, нормальные, сбалансированные и несбалансированные, имеющие как транслоцированную, так и нормальную хромосому 21. В комбинации с нормальной гаметой это может приводить к зачатию ребенка с транслокационным синдромом Дауна. Теоретически эти три типа гамет производятся в равных количествах, таким образом, теоретический риск ребенка с синдромом Дауна должен быть 1 к 3. Тем не менее расширенные популяционные исследования показали, что несбалансированные хромосомные наборы появляются только у 10-15% потомства матерей и только у нескольких процентов потомства отцов, несущих транслокации, включающие хромосому 21. Транслокация 21q21q при синдроме Дауна. Хромосомная транслокация 21q21q — хромосома, сформированная из двух длинных плеч хромосомы 21; бывает у нескольких процентов пациентов с синдромом Дауна. Считают, что они появляются как изохромосомы, а не робертсоновские транслокации. Большинство таких случаев возникают постзиготически, соответственно, риск повторения низкий. Тем не менее особенно важно убедиться, не является ли родитель носителем (возможно, мозаичным) данной транслокации, поскольку все гаметы носителя такой хромосомы должны также содержать 21q21q хромосому, с двойной дозой генетического материала хромосомы 21, или не иметь хромосомы 21 совсем. Потенциальное потомство, следовательно, неизбежно имеет или синдром Дауна, или нежизнеспособную моносомию 21. Мозаичные носители имеют повышенный риск повторения, таким образом, пренатальная диагностика необходима при всех последующих беременностях. Мозаичный синдром Дауна. Около 2% пациентов с синдромом Дауна — мозаики, обычно с популяциями нормальных клеток и с трисомией 21. Фенотип может быть мягче, чем при типичной трисомии 21. Вообще существует широкая изменчивость в фенотипах мозаичных пациентов, вероятно, отражая различные пропорции трисомных клеток у эмбриона на ранних стадиях развития. Возможно, пациенты с установленным мозаичным синдромом Дауна отражают только клинически более серьезные случаи, поскольку в легких случаях кариотипирование менее вероятно. Частичная трисомия 21 при синдроме Дауна. Очень редко синдром Дауна диагностируют у пациентов, имеющих трисомию только по части длинного плеча хромосомы 21, и еще реже выявляют пациентов с синдромом Дауна без цитогенетически видимой хромосомной аномалии. Такие случаи представляют определенный интерес, поскольку могут указывать, какая область хромосомы 21, вероятно, ответственна за специфические компоненты фенотипа синдрома Дауна и какие области могут утраиваться, не вызывая фенотипических проявлений. Хотя хромосома 21 содержит только несколько сотен генов, попытки согласовывать тройную дозу специфических генов со специфическими аспектами фенотипа синдрома Дауна пока имеют ограниченный успех. Наиболее примечательной стала идентификация области, критической для пороков сердца, наблюдаемых примерно у 40% пациентов с синдромом Дауна. Поиск конкретных генов, существенных для проявления фенотипа синдрома Дауна, среди случайно находящихся рядом с ними в хромосоме 21, — главная задача современных исследований, особенно на мышах в качестве модели. Потенциально перспективное направление — исследование генно-инженерных мышей с дополнительной дозой генов из хромосомы 21 человека (или даже с полной копией хромосомы 21). Такие мыши могут проявлять фенотипические аномалии в поведении, функциях мозга и формировании сердца. – Также рекомендуем “Причины синдрома Дауна. Риск рождения ребенка с трисомией 21″ Оглавление темы “Хромосомные аномалии”:
|
Источник